現在の研究テーマ
1.核膜融合のメカニズム
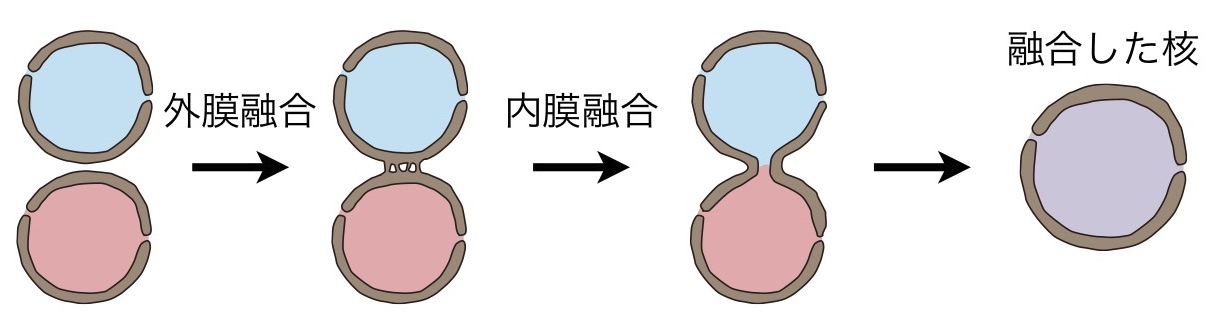
酵母や植物の有性生殖過程では、核膜の融合によって核融合が行われます。私たちは、有性生殖過程の核融合過程に欠損を示す出芽酵母とシロイヌナズナの変異株を利用した分子遺伝学的アプローチで、核膜融合にあずかるタンパク質の同定を進めてきました。その結果、酵母と植物という進化的に離れた生物の間で、有性生殖過程の核膜融合の分子機構が保存されていることがわかりました。たとえば、小胞体の分子シャペロンHsp70や核内膜のSUNタンパク質が、これらの核融合過程で共通に機能することを示してきました。また、これらタンパク質に関する変異株の解析の結果、核膜融合は核外膜の融合と核内膜の融合という2回の膜融合によって進行し、これらの膜融合過程は異なるタンパク質によって制御されていることもわかってきました(図)。卵細胞などの生殖細胞には、有性生殖過程で核膜融合を促進する特別な仕組みがあると考えられます。私たちは最近、有性生殖過程特異的に発現するシロイヌナズナの核膜タンパク質GEX1を同定しました。GEX1はシロイヌナズナの有性生殖過程で観察される3回の核融合すべてに必要であること、GEX1が核膜融合の過程に必要であることを明らかにしました。また、GEX1は卵細胞と中央細胞といった生殖細胞特異的に発現する核膜タンパク質であることを明らかにしました。
私たちは現在、核膜融合で機能する新たなタンパク質の同定を目指しています。このため、出芽酵母やモデル植物シロイヌナズナの変異株を用いた分子遺伝学的アプローチに加えて、ライブイメージング解析、配偶体特異的遺伝子発現誘導系やケミカルバイオロジーなどの新たな手法を用いて研究を行っています。
2.受精とカップルした精核融合の意義
被子植物の重複受精では、花粉管から放出された2つの精細胞が、雌性配偶体の卵細胞と中央細胞それぞれと融合して胚および胚乳ができます。精細胞の核(精核)は、細胞融合の後すぐに卵細胞または中央細胞の核と融合し、胚または胚乳の核ができます。シロイヌナズナの核融合変異株は、受精の過程に欠損を示しませんが、胚乳形成が異常となるため種子が死にます。私たちは、シロイヌナズナの核融合変異株の胚乳形成過程のライブイメージング解析を行い、中央細胞における受精とカップルした精核の融合が、精細胞由来の遺伝子の発現と胚乳形成に重要な役割をはたしていることを明らかにしました。現在は、卵細胞における精核融合についても解析を進めています。3.核融合機構の多様性−様々な生物で核融合のしくみに違いはあるのか
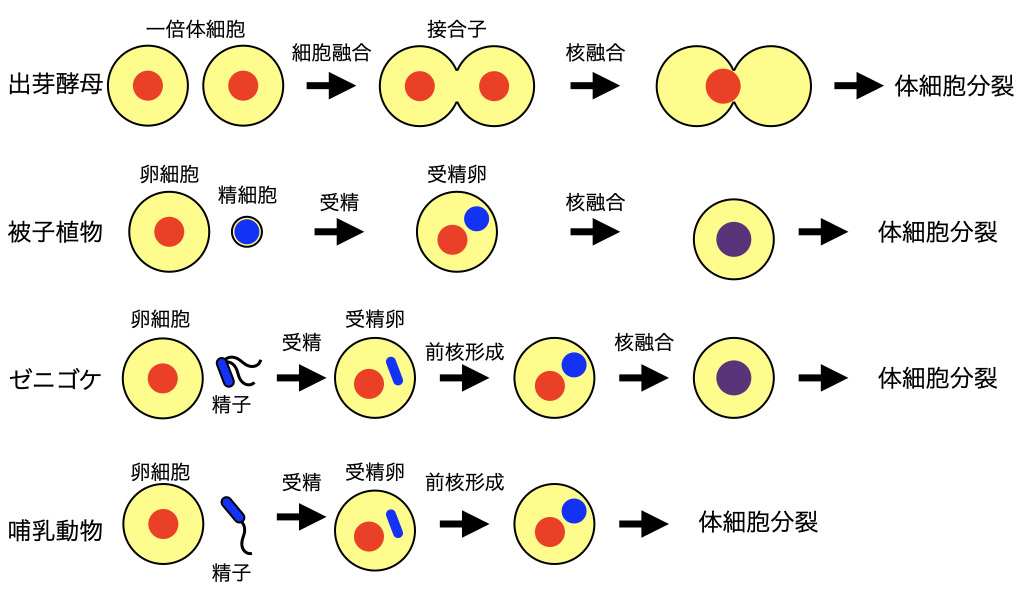
出芽酵母接合時の核融合は、同じ大きさ・状態の核同士の融合であるのに対して、被子植物重複受精の際の精核融合は、大きさや状態が異なる核同士(例えば精核と卵細胞核)の融合です。しかし、これらの核膜融合で機能するタンパク質には共通性が見られます。被子植物の受精過程では、花粉管が運んできた精細胞によって受精が行われます。一方で、動物や、植物でもコケ・シダ類などの生殖過程では、卵と精子が受精します。これらの生物では、受精後に精子の核は雄性前核へと変化し、卵由来の核(雌性前核)と融合します。雌雄前核が融合して接合子の核ができた後に胚発生が始まる生物も多いです。一方で哺乳動物では、雌雄前核の融合が最初の体細胞分裂と同じタイミングでおこるので、細胞分裂時の核膜崩壊によって雌雄の核が融合します。このような核融合でも、酵母や被子植物の場合と同様なタンパク質が機能しているのでしょうか。われわれは卵と精子が受精する生物における核融合機構の解明を目標に、コケ植物のゼニゴケを用いた研究も進めています。
参考文献
- Nishikawa and Endo, J. Biol. Chem. 1997 DOI
- Nishikawa et al., J. Biol. Chem. 2003 DOI
- Nishikawa et al., Genes Cells 2008 DOI
- Maruyama et al., Proc. Natl. Acad. Sci. USA 2010 DOI
- Maruyama et al., Plant Cell Physiol. 2014 DOI
- Hwang et al., Plant Cell Physiol. 2019 DOI
- Maruyama et al., Plant Cell Physiol. 2020 DOI
- Nishikawa et al., Front. Plant Sci. 2020 DOI